
 Les derniers travaux de Lynn Margulis portaient sur l’origine du noyau. C’est là une étape cruciale dans la mesure où elle marque la naissance des eucaryotes, distincts des procaryotes, qui évolueront plus tard en plantes, animaux, champignons.
Les derniers travaux de Lynn Margulis portaient sur l’origine du noyau. C’est là une étape cruciale dans la mesure où elle marque la naissance des eucaryotes, distincts des procaryotes, qui évolueront plus tard en plantes, animaux, champignons.
Autrement dit, cette étape marque le déroulement de la première symbiose.
Lynn Margulis s’est donc intéressée au « Dernier Ancêtre Eucaryote Commun » (en anglais, LECA, pour last eukaryotic common ancestor) qui correspond à l’acquisition de la motilité cytosquelettique.
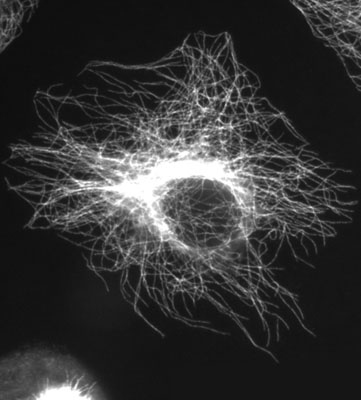
Le cytosquelette est composé de sortes de filaments qui se renouvellent constamment et assurent le maintien ou la transformation de la forme de la cellule, mais aussi de nombreuses fonctions comme le transport des chromosomes lors de la division de la cellule, ou mitose.
La mitose est également d’une grande importance dans la recherche du Dernier Ancêtre Eucaryote Commun, puisque la mitose n’existe que chez les eucaryotes. En effet, chez les procaryotes, la division cellulaire se nomme la fission binaire et elle ne fait intervenir aucune motilité cytosquelettique.
La mitose, ou division cellulaire asexuée, se déroule en six phases : l’interphase, la prophase, la prométaphase, la métaphase, l’anaphase et la télophase.
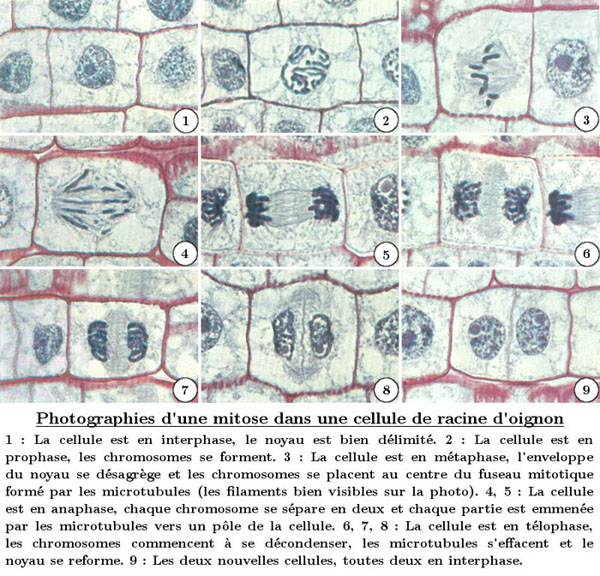
Comme on le voit sur les photographies ci-dessus, un fuseau mitotique se déploie pendant la mitose pour permettre la répartition des chromosomes. Ce fuseau est composé de microtubules, des sortes petits tubes dont nous avions déjà parlé dans l’article précédent sur les ondulipodes et de leur structure particulière en 9+2.
 Parmi tous les filaments dont est composé le cytosquelette, Lynn Margulis va donc s’intéresser à ce type de ces filaments en particulier : les microtubules. Or, il se trouve que la particularité de certains spirochètes est de contenir de la tubuline, la protéine qui forme les microtubules.
Parmi tous les filaments dont est composé le cytosquelette, Lynn Margulis va donc s’intéresser à ce type de ces filaments en particulier : les microtubules. Or, il se trouve que la particularité de certains spirochètes est de contenir de la tubuline, la protéine qui forme les microtubules.
Lynn Margulis va alors émettre l’hypothèse que l’origine d’une partie de la motilité cytosquelettique provient d’une symbiose avec des spirochètes. Plus précisément, elle présume que le noyau et le centrosome, parfois appelé système organisateur des microtubules, sont les évolutions de la symbiose entre une archéobactérie et un spirochète.
Pour appuyer cette hypothèse, Lynn Margulis fait appel au caryomastigonte, un ensemble décrit pour la première fois en 1915 mais qui n’avait jamais été utilisé dans le cadre de l’évolution. Le caryomastigonte est un ensemble constitué d’ondulipodes connectés à un noyau par un connecteur nucléaire (le rhizoplaste).
Le croquis ci-dessous montre comment le Dernier Ancêtre Eucaryote Commun a évolué à partir de la symbiose entre un micro-organisme Thermoplasma et un spirochète.
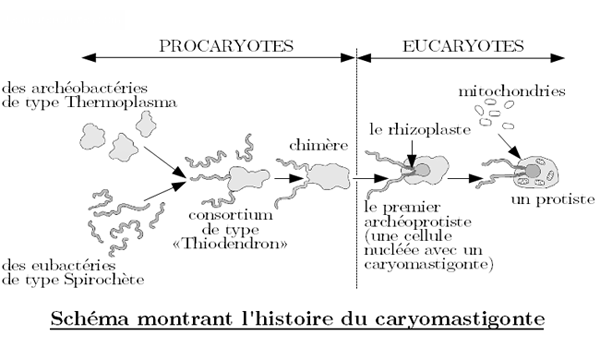
Selon cette hypothèse, l’apparition du noyau et de la mitose (et donc des structures qui lui sont liées) se sont donc réalisées en même temps. Et cette apparition daterait du Protérozoïque, qui s’étend d’il y a 2500 à 541 millions d’années.
Pendant longtemps, la façon dont le centrosome (le système organisateur des microtubules) a émergé est restée une énigme. En 1987, Lynn Margulis disait même que c’était une « énigme centrale de la biologie cellulaire ». L’hypothèse basée sur la symbiose de deux bactéries menant au Dernier Ancêtre Eucaryote Commun vient donc combler un trou laissé béant depuis de nombreuses années.
Et dans cette hypothèse, le caryomastigonte a un statut particulier : c’est ce système qui marque véritablement la naissance des eucaryotes, de « l’individualité complexe » des eucaryotes comme le dit Lynn Margulis.